I’ve had this blog a year and a half now and although I’ve had a good time, I’ve never really wrote about anything I’m actually (somewhat) knowledgeable. So I think I’ll do a few posts on leatherback turtles. I’m no turtle expert but I did spend a couple years working on their migration and movement. My first first-author paper just got officially published so I’ll start with that. <ego>It made the cover of Behavioral Ecology</ego> which isn’t really a big deal but I thought it was kind of cool. The paper was about leatherback turtles migrating from their summer feeding grounds near Nova Scotia to head down to wintering and nesting areas in the south. (If you don’t feel like reading all this, you could just skip to the pretty cool {at least to me} movie down at the bottom).
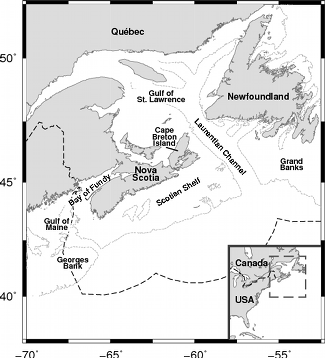
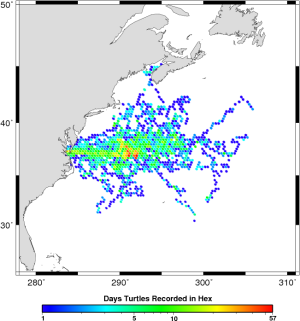
First a little background. If you don’t know much about Nova Scotian geography, don’t feel too bad a lot of my relatives in Michigan were surprised when I told them I was in Canada but still south of them. Here’s a map for orientation. Leatherback turtles come up to Nova Scotian waters to eat jellyfish in the summer and then head back south to nest, mate and presumably avoid the cold waters. Leatherback turtles are pretty cool animals and I’ll probably do another post with more details on them later. For now, you just need to know they’re big animals reaching more than half a ton and their size, blubber and counter-current heat exchange allow them to maintain a high body temperature in cold waters almost like a warm-blooded animal.
So back to the paper, my coauthor Mike James has tagged a whole bunch of turtles now and we were curious if we could see any pattern in when they begin their migration. One problem was that unlike migratory birds it’s pretty hard to define when a turtle begins migrating. Luckily the state space models of Ian Jonsen provide a method to estimate turtle behavior as either foraging or transiting. By finding the last foraging in a season, we could have a pretty good idea of when the turtles began their migration.
Once we figured out when the turtles began their migration, we tried to figure out what might be triggering it. We guessed it would have something to do with colder temperatures or declining prey abundance. I could get temperature from satellite data but there isn’t an easy way to measure jellyfish (leatherback food) abundance. As a rough proxy, we used chlorophyll estimates from satellite images. As a side note, it’s really cool that NASA provides their data for free. Just to cover as many bases as possible, we threw position, day length, the North Atlantic Oscillation index, water depth and the sex and size of the turtle into the mix and stuck it all into a stats model. After a bit of calculating, the model came back predicting that the position of the turtle and temperature and chlorophyll of the water appear to correlate with departures.
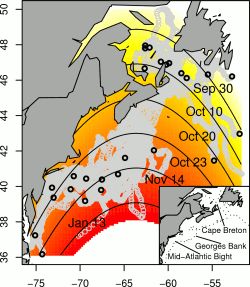
The biggest factor for predicting departure was the position of the turtle. Northern turtles appear to leave earlier than southern turtles and turtles around the longitude of Georges Bank (abundant shelf ecosystem) and Nova Scotia (the study area and lots of jellyfish) stay longer. The contour plot to the right shows the relative probability of departure from low (red) to high (yellow). The contour lines and dates show when we would expect 50% of the turtles in a region to depart. Knowing these dates could be pretty useful for conservation since regulations could be lifted after their departure.
Northern turtles leaving earlier was pretty much what we expected since northern waters get colder sooner. So it was pretty surprising when the model also predicted that turtles are actually more likely to leave when the water is warmer and greener. Now I’m not sure exactly why this is but one possible guess is that warmer chlorophyll-rich waters provide more nourishment sooner and allow turtles to head south earlier. Another possible explanation is that jellyfish population might decrease earlier since many jellyfish die off after reproducing and warmer waters and increased prey allow jellyfish to reproduce sooner. Of course, a final possibility is that the correlation is just a funny coincidence in the data but hopefully this is unlikely.
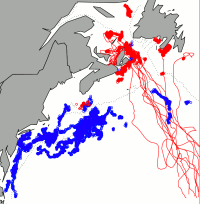
Since turtles didn’t appear to be leaving when the water cooled off, why were they still leaving early from the north? One guess might be that turtles leave earlier from the north if they have farther to swim to their southern nesting grounds but the difference from the southern and northern foraging areas is only about 500 km. Since leatherbacks can easily swim 2 km/hr, they could cover this distance in 10 days or less but we observed differences of more than a month. Without temperature or distance to explain it, the disparity could possibly be related to some difference in habitat quality. Unfortunately this will remain just a hypothesis until we get a lot more data on jellyfish distribution and feeding rates. It is interesting to note that although many leatherbacks are still foraging in southern waters (blue dots in the figure to the right), no northern foraging turtle (red dots) has been observed moving into southern foraging grounds late in the season (red tracks).
Now that you made it through all that text, here’s a video I made of the turtles foraging, transiting and migrating (higher quality here). By the way, it costs quite a bit of work and money to get a transmitter on a leatherback so each one of those points is a good bit of work done by my coauthor Dr. James.
A couple extra interesting things to watch for are turtles hitting the Gulf Stream east of the continental shelf and being swept to the northeast and turtles getting stuck in the Bay of St. Lawrence (there used to be a channel between Cape Breton Island and mainland Nova Scotia but it was recently filled in with a causeway). See the map above if you need help finding those. Anyway, that movie’s pretty much the highlight of my thesis. I must have seen it several dozen times already but I still get a kick out of watching the turtles swim around.
Reference
Sherrill-Mix, S.A., James, M.C., Myers, R.A. (2007). Migration cues and timing in leatherback sea turtles. Behavioral Ecology, 19(2), 231-236. DOI: 10.1093/beheco/arm104